







In ogni battaglia se conosci il nemico, e te
stesso, hai già iniziato a vincere e oggi 13 Marzo 2020, in pieno periodo Coronavirus
– Covid-19 – è doveroso conoscere il meccanismo di difesa del nostro sistema immunitario,
perché NOI non siamo inermi, anzi, possediamo delle robuste
difese che ci aiutano a fronteggiare ogni tipo di aggressione con cui
entriamo in contatto…direi quindi di iniziare, conscendo i meccanismi di difesa
contro i virus che il nostro
------------
“Come il sistema immunitario si è evoluto per
combattere le infezioni così gli
agenti patogeni si sono evoluti per sfuggirlo. La forma e la struttura degli
agenti patogeni e del sistema immunitario sono il prodotto di questa costante
battaglia per la sopravvivenza. Durante questo processo di coevoluzione, sia
gli agenti patogeni sia il sistema immunitario hanno sviluppato i propri punti
di forza, ma anche le proprie debolezze. Questa competizione ha prodotto un
sistema con molteplici livelli di difesa. Gli agenti patogeni si scontrano
inizialmente con le difese innate non adatta bili, poi con quelle specifiche. Questi
due meccanismi si fondono armoniosamente e diversi processi si svolgono
parallelamente, in modo sia indipendente sia cooperativo. Molte delle tipologie
di interazione tra sistema immunitario e agente patogeno sono attualmente
conosciute a livello molecolare. Malgrado il raggiungimento di questi
traguardi, le infezioni costituiscono ancora la maggiore minaccia per la salute
nel mondo. La sfida per il futuro consisterà nell'applicare le nostre
conoscenze in campo immunologico per debellare i continui attacchi dei germi
patogeni.
Introduzione
Durante tutta la vita, il sistema immunitario ci
difende dai continui attacchi di agenti patogeni potenzialmente letali, mentre
tollera microbi che non rappresentano un'immediata minaccia. I microbi che si
sviluppano sulla cute e nell'intestino superano in rapporto di circa 10:1 le
nostre cellule. Si è stimato che il sistema immunitario affronti ed elimini,
solo nel primo anno di vita, in media nove infezioni virali a livello
respiratorio (Denny, 1995), ciascuna delle quali genera un nuovo insieme di
antigeni che devono essere riconosciuti dallo stesso sistema. In ogni
successivo anno di vita, il sistema immunitario incontra numerosi nuovi
antigeni.
I meccanismi effettori che eliminano le infezioni
costituiscono un'intricata rete, i cui singoli componenti sono tra loro
interconnessi. In tabella (tab. I) sono indicate le principali
componenti di questo sistema sequenziale di difese immunitarie (fig. 1). Sebbene la rete sia piuttosto
complessa, si conoscono attualmente con notevole dettaglio le basi molecolari
del funzionamento delle singole componenti.
La via preferenziale di ingresso della maggior parte
degli agenti patogeni virali è costituita dalle mucose. La minaccia delle
infezioni respiratorie e intestinali alla salute mondiale è in un certo senso
inevitabile. Respirare, bere, mangiare e avere rapporti sessuali sono attività
ineliminabili; queste funzioni necessitano di scambi di materiali con
l'ambiente e con altri ospiti. Qualsiasi agente patogeno ben adattato è spinto
a sfruttare queste funzioni per guadagnarsi l'accesso all'ambiente interno
dell'organismo.
La cute e le mucose non sono solo i luoghi ideali di
invasione, ma sono anche punti strategici per diffondere gli agenti patogeni ad
altri ospiti. L'irritazione della mucosa nasale causata dal comune virus del
raffreddore provoca la secrezione di muco, che contiene abbondante virus
infettivo. Starnuti e tosse causano l'espulsione delle secrezioni. Particelle
di muco si depositano sulle mucose dei nuovi ospiti. Il ciclo dell'infezione
non richiede che il virus si diffonda in altri tessuti al di fuori del tratto
respiratorio.
La trasmissione può in realtà essere più 'efficace' se
le condizioni della persona infetta non sono così gravi da dover evitare i
contatti sociali. Gli agenti che causano la diarrea e il vomito garantiscono la
loro diffusione tramite questi effetti, che raramente sono letali per l'ospite.
Si può sostenere che gli agenti patogeni che causano il comune raffreddore e
forme acute gastrointestinali hanno raggiunto uno stadio ideale di evoluzione
stabile. Aumentare o diminuire la patogenicità sarebbe uno svantaggio per
l'aggressore. Gli agenti patogeni che causano elevata mortalità possono forse
essere considerati 'intrusi' instabili, che ancora devono evolversi fino allo
stato di equilibrio con l'ospite.
Le stime dell'Organizzazione mondiale della sanità
indicano che circa 14 milioni di persone muoiono ogni anno di infezioni
trasmesse attraverso le vie respiratorie, a confronto con i 5 milioni di
decessi per malattie trasmesse attraverso la via gastrointestinale, che
colpiscono per lo più i bambini. Le infezioni virali dei bambini sono di
importanza fondamentale nel loro sviluppo e nell'andamento delle malattie
nell'adulto. Si spera che ulteriori progressi nella comprensione delle difese
contro le malattie infettive rendano tali patologie soggette a prevenzione e
trattamento, riducendo in tal modo anche il loro costo per la salute umana.
Difese a livello di superficie
l virus hanno bisogno di cellule viventi per replicarsi
ed è significativo che quasi tutte le cellule visibili sulla cute siano morte.
l papillomavirus hanno bisogno di una lacerazione della cute per formare una
verruca; anche l'HlV non riesce a infettare attraverso la cute intatta. Sebbene
lo strato superficiale del tratto respiratorio e di quello gastrointestinaIe
contengano cellule vive, la maggior parte di esse muore e viene eliminata dall'
organismo in 24 ore. Questo 'muro di cadaveri' è senza dubbio la più importante
singola difesa contro le infezioni invasive. Le cellule morte, tuttavia, non
sono le sole difese di superficie. Batteri e funghi non patogeni, cioè la
normale flora, ricoprono la cute, il tratto aereo superiore e il tratto
digestivo, competendo con batteri potenzialmente nocivi per i siti di adesione
e per le sostanze nutritive. Riducendo la densità di questa flora normale con
gli antibiotici si può aumentare il potenziale invasivo degli agenti microbici
non sensibili ai trattamenti. La competizione per una posizione offre una
barriera importante alle infezioni invasive.
Nel tratto respiratorio vi è un costante flusso di muco
dalle regioni profonde dei polmoni e dal naso verso la faringe, dove il muco è
normalmente deglutito o in certi casi eliminato. Questo velo di muco comprende
isole di spesso gel in un mare di sol. Il muco contiene enzimi antibatterici e
antivirali, come illisozima, presente nella saliva, nel sudore e nelle lacrime.
Inoltre le mucoproteine che contengono acidi sialici si adsorbono su virus come
quello dell'influenza A, riducendo la capacità dei virus stessi di aderire ai
recettori delle mucose. La cute, invece, produce secrezioni sebacee contenenti
acidi grassi, che sono potenti agenti antimicrobici. Nell'intestino,
l'alternanza di pH basso e alto e la peristalsi che smuove il materiale dallo
stomaco all'intestino tenue, le proprietà detergenti ed enzimatiche della bile e
delle altre peptidasi intestinali eliminano gran parte degli agenti infettivi
che entrano nel tratto intestinale. Sulle mucose si è evoluto un sistema di
difesa complesso, esteso e altamente specializzato. La presenza di materiali
estranei sulla superficie di tali mucose necessita di un'attenta regolazione da
parte delle risposte immunitarie per evitare danni ai tessuti vicini. La
funzione di queste difese di superficie consiste nella protezione contro
l'invasione da parte di potenziali agenti patogeni, tollerando però materiali
non patogeni. Al contrario, il sistema immunitario interno funziona normalmente
in un ambiente sterile e reagisce in modo più vigoroso alle sostanze
antigeniche. Il successo dell'eliminazione degli antigeni dalle mucose,
ottenuto senza danneggiare i tessuti vicini, dipende da risposte immunitarie
bilanciate. Se i meccanismi immunitari non infiammatori falliscono nell'
eliminazione dell' antigene, prevarranno meccanismi pro infiammatori che
causeranno un danno. Questa alterazione immunitaria è responsabile di alcune
manifestazioni cliniche che si verificano durante l'infezione virale.
Risposte immunitarie innate
Macrofagi
l macrofagi (letteralmente, grosse cellule mangiatrici)
sono una componente chiave del sistema immunitario innato. Essi inglobano i
virus e altri agenti patogeni che sono stati neutralizzati dagli anticorpi
circolanti e che devono essere distrutti senza danneggiare i tessuti
circostanti. l macrofagi derivano da quelle cellule del midollo osseo che
diventano monociti del sangue. Questi si differenziano in macrofagi nei
polmoni, nei linfonodi, nella milza, nella cute, nel sistema nervoso centrale
(dove formano la microglia) e nel fegato (dove costituiscono le cellule di
Kupffer). Sebbene la loro funzione primaria sia inglobare la materia
particolata, l'ingestione è più efficace se la particella è ricoperta dagli
anticorpi o dal complemento, che hanno entrambi la funzione di opsonizzare la
particella. Si può paragonare questo processo di opsonizzazione all'aggiunta di
salse o sughi ai cibi per renderli più gustosi e appetibili. Dopo aver
inglobato la particella, il macrofago digerisce il contenuto della vescicola
endocitata. l prodotti di digestione della particella vengono trasportati alla
superficie cellulare in associazione con il complesso maggiore di
istocompatibilità (MHC, Major Histocompatibility Complex). Se questo frammento
peptidico non è riconosciuto come proprio dell' ospite, vengono attivati i
linfociti T helper (v. oltre).
Il processo di ingestione inizia con il legame del
materiale opsonizzato al recettore del frammento cristallizzabile sulla
membrana dei macrofagi e continua con l' endocitosi in una vescicola interna,
chiamata fagosoma. Quest'ultimo si fonde con una vescicola, contenente lisozima
e altri enzimi digestivi, chiamata liso soma; la vescicola che risulta dalla
fusione si chiama fagolisosoma. Una combinazione di pH basso, mantenuto dalle
pompe ioniche, di idrolasi acide, di perossido di idrogeno, di radicali
reattivi dell'ossigeno e di ossido d'azoto è in grado di demolire il contenuto
del fagoliso soma. Talvolta la particella da distruggere è troppo grossa per
essere fagocitata, come nel caso di un parassita. Questo provoca il rilascio
degli enzimi lisosomiali nel mezzo extracellulare, con la possibilità di
causare danni localizzati alle cellule dell'ospite.
I macrofagi sono predominanti nei granulomi, processi
infiammatori in cui vari tipi di cellule effettrici (monociti, linfociti,
cellule polimorfonucleate), attratte dai fattori chemiotattici rilasciati dopo
l'invasione del tessuto da parte di materiali estranei particolati (inclusi
agenti patogeni), formano una barriera attorno alla particella estranea. Esempi
tipici di questo fenomeno si osservano nella tubercolosi, dove micobatteri (gli
agenti che causano la tubercolosi o la lebbra) possono sopravvivere per decenni
all'interno di un granuloma.
Molti agenti microbici importanti hanno sviluppato
sistemi per evitare gli effetti antimicrobici dei macrofagi. Alcuni virus,
specialmente dengue e HIV, si sviluppano effettivamente dentro i macrofagi e
possono usare anticorpi antivirali per attivare la loro capacità di infettare i
macrofagi stessi, che divengono inefficaci e funzionano persino da 'cavallo di
Troia', trasportando l'infezione in altre regioni del corpo. Alcuni agenti
patogeni non virali (per esempio, Leishmania e molti micobatteri) sono ospiti
intracellulari obbligati e riescono a replicarsi soltanto all'interno della
cellula. I macrofagi sono efficienti non solo nel digerire antigeni e nel
presentare componenti estranee, ma anche nel produrre alcune citochine, fra cui
le interleuchine l e 6 (IL-l e IL-6). Un macrofago infetto allerta perciò le
cellule circostanti, inclusi i linfociti B, della presenza di materiale
estraneo potenzialmente infettivo.
Cellule natural killer e interferoni
Una seconda componente della risposta immunitaria
innata è costituita dal sistema delle cellule natural killer (NK) che di solito
è collegato con l'azione degli interferoni (fig. 2).
L'interferone α/β, o interferone di tipo I, è prodotto
da una grande varietà di cellule in seguito all'infezione virale e attiva la
citotossicità mediata dalle cellule NK tramite il legame a specifici recettori.
Alcuni virus sono in grado di produrre una molecola solubile analoga al
recettore dell'interferone di tipo I che inibisce l'effetto antivirale
dell'interferone stesso, competendo con i normali meccanismi di attivazione.
L'esistenza di questo meccanismo di evasione virale suggerisce un ruolo
importante dell'interferone nella resistenza ai virus. Oltre che dalle cellule
NK, l'interferone γ è rilasciato dai linfociti T attivati; esso induce anche
uno stato antivirale, per esempio inducendo nel topo il gene Mx che genera
resistenza al virus influenzale, e stimola il processamento e la presentazione
dell'antigene in modo simile all'interferone di tipo I. L'interferone γ è
prodotto dalle cellule T che si infiltrano nei gangli neuronali infettati dal
virus Herpes simplex e si ritiene possa essere coinvolto nel passaggio dalla
fase litica al ciclo di replicazione latente del virus (v. oltre).
Le cellule NK differiscono dalle cellule T citotossiche
in quanto non si differenziano nel timo. Il meccanismo attraverso cui le
cellule NK riconoscono ed eliminano gli agenti patogeni è rimasto oscuro fino a
tempi molto recenti (Biron, 1997). Le cellule NK sono reclutate entro i primi
due giorni dall'infezione e si dividono e si differenziano localmente. Esse
hanno importanti effetti antivirali, in parte perché uccidono le cellule
infettate attraverso la formazione di granuli citolitici e in parte perché
producono interferoni α, β e γ. Molti progressi in questo campo hanno riacceso
l'interesse per le cellule NK, in particolare quelli riguardanti la descrizione
di numerosi recettori esposti sulla superficie delle cellule NK e di potenziali
cellule bersaglio che regolano l'attività delle cellule NK stesse. La prima interazione
di questo tipo avviene tra l'MHC di classe I sulle normali cellule bersaglio e
i recettori inibitori dei killer (KlR, Killer Inhibitory Receptor) sulle
cellule NK (fig. 3). Il coinvolgimento delle molecole di
classe I manda un segnale negativo alle cellule NK, inibendo la loro azione
litica. L'assenza o l'alterazione di molecole di classe I previene la
stimolazione di questo segnale inibitorio. La cellula NK viene quindi attivata
da segnali trasmessi attraverso un secondo recettore e ciò provoca il rilascio
del contenuto della cellula NK. Il rilascio direzionale di granuli provoca
apoptosi nella cellula bersaglio. Qualsiasi cellula contenente un virus che
diminuisca l'espressione delle molecole di classe I è soggetta a essere uccisa
da questo meccanismo (v. il saggio di E.I.H.I. Wiertz, nel II volume).
Alcuni virus, come per esempio il citomegalovirus (CMV)
umano e murino, cercano di sabotare questo meccanismo codificando molecole
omologhe all'MHC di classe I. Inoltre, le cellule normali non infettate
rispondono al rilascio locale di interferone aumentando l'espressione della
classe I. Il fenomeno rende queste cellule, normalmente sensibili, resistenti
all'uccisione da parte delle cellule NK. Se le cellule sono infettate, questa
aumentata espressione può non avvenire, rendendo le cellule più suscettibili
all'uccisione mediata da cellule NK. Questo è uno degli esempi dell'azione dei
recettori inibitori dei killer, ma le cellule NK possono provare una varietà di
KlR prima di raggiungere un punto definitivo e svuotare i granuli citotossici.
Una volta che le risposte specifiche locali delle cellule T sono attivate,
meccanismi aggiuntivi inibiscono le funzioni delle cellule NK. L'attività di
queste cellule potenzialmente dannose è perciò di breve durata, ma copre
l'intervallo tra l'inizio dell'infezione e lo sviluppo dell'immunità specifica.
Risposte immunitarie acquisite
Mentre le risposte immunitarie innate vengono innescate
precocemente e hanno scarsa o nessuna specificità per il particolare agente
patogeno, quelle acquisite si verificano più tardi nel tempo ma sono altamente
specifiche. llinfociti specifici per l'antigene proliferano e si differenziano
durante questo intervallo di tempo, raggiungendo valori che sono sufficienti a
costituire una difesa efficace. Se si è incontrato un agente patogeno identico
o simile in precedenza, il ritardo si riduce e la potenza della risposta
immunitaria è fortemente rafforzata; questo fenomeno viene definito memoria
immunitaria. Sulle mucose, le cellule linfoidi si concentrano in aggregati
specializzati chiamati MALT (Mucosal Associated Lymphoid Tissue, tessuto
linfoide associato alla mucosa). Quando queste cellule migrano,
preferenzialmente tornano indietro, proprio nelle aree mucose. Questo processo
di homing (localizzazione) è dovuto all'espressione di recettori di superficie
che riconoscono in modo specifico le addressine vascolari, presenti
sull'endotelio di venule specializzate dei tessuti linfoidi, le HEV (High
Endothelial Venules, venule con endotelio alto).
Linfociti B
Uno dei principali tipi cellulari coinvolti
nell'immunità acquisita è costituito dai linfociti B. Le cellule B sono così
chiamate perché sono state studiate nella borsa di Fabrizio del pollo. In
questa specie, l'asportazione della borsa provoca l'eliminazione delle cellule
B e della produzione di anticorpi durante lo sviluppo. Le cellule B mature
portano in superficie le immunoglobuline, che agiscono come recettori dell'
antigene, e sono distribuite dal sangue e dalla linfa ai tessuti linfoidi, in
particolare ai linfonodi e alla milza. Non esiste un equivalente anatomico
della borsa di Fabrizio nei mammiferi, dove lo sviluppo delle cellule B inizia
nel midollo osseo dalle cellule staminali che derivano originariamente dal
fegato. Queste cellule formano un lignaggio attraverso le cellule pro-B, le
cellule pre-B, fino alle cellule B vere e proprie e infine alle plasmacellule
(v. il saggio di F. Melchers, Lo
sviluppo dei linfociti). Per progredire in questo processo di
maturazione, le cellule B rispondono a stimoli di sviluppo, in modo particolare
a lL-7 prodotta dalle cellule stromali nel midollo osseo. Durante l'ontogenesi,
i geni che codificano le molecole di anticorpi vengono ricombinati per produrre
catene pesanti e leggere uniche che costituiscono il recettore specifico per
l'antigene (v. il saggio di C. Rada, nel II volume). La plasmacellula
rappresenta lo stadio finale di sviluppo, durante il quale sono se crete
abbondanti quantità di anticorpi. Esistono due tipi principali di risposte da
parte delle cellule B, quella dipendente dalle cellule T e quella indipendente
dalle cellule T. È stato proposto che la via T-indipendente sia diretta
soprattutto contro gli antigeni ripetitivi, come quelli presenti sulla
superficie di molti agenti microbici, e che questa via sia responsabile
dell'induzione di potenti risposte antivirali. Tali risposte sono più rapide,
non richiedono aiuti, di preferenza inducono risposte neutralizzanti e molto
durature, soprattutto se confrontate con le risposte T -dipendenti (Bachmann e
Zinkemagel, 1996). Al contrario, le risposte T -dipendenti, come quelle verso
le proteine coniugate agli apteni o agli eritrociti di pecora, dipendono dalla
costimolazione delle cellule B di molecole accessorie, che avviene grazie alle
interazioni che si stabiliscono tra gli anticorpi monoclonali B7.1 o B7.2, o
entrambi, sui linfociti e il CD28 sulla superficie dei linfociti T helper, o
tra CD40 e il suo ligando. L'interazione con i linfociti B induce inoltre i
linfociti T helper a rilasciare citochine, che inducono la proliferazione, la
produzione di anticorpi e lo switch (commutazione) isotipico dei linfociti B.
Le immunoglobuline 'saggiano' tutte le proteine con le
quali vengono in contatto e formano legami stabili solo con molecole la cui
superficie mostra caratteristiche chimicofisiche, come forma, distribuzione di
cariche e idrofobicità, che esprimono la maggiore complementarità per
l'immunoglobulina (v. il saggio di R.A. Mariuzza, Riconoscimento
dell'antigene da parte dei recettori del sistema immunitario).
Quando la corrispondenza spaziale è esatta, l'immunoglobulina resta attaccata
sufficientemente a lungo per innescare altre risposte. Ciò significa che devono
esistere tante immunoglobuline sufficientemente differenti per riconoscere
quasi tutte le proteine patogene esistenti e potenziali. Nello stesso tempo, si
deve evitare il riconoscimento delle molecole proprie dell'organismo.
Individuare il modo in cui sono prodotte le diverse immunoglobuline è stato uno
dei principali enigmi dell'immunologia, risolto solo pochi anni or sono. La
generazione della diversità è raggiunta combinando molti diversi segmenti genici
codificanti polipeptidi che riarrangiano durante lo sviluppo per produrre
catene ibride leggere e pesanti (v. i saggi di G.J.V. Nossal e di C. Rada, nel
II volume). Nella linea germinale esistono molti elementi variabili che possono
ricombinarsi con una notevole percentuale di 'imprecisione'. Le mutazioni
puntiformi sono frequenti e diverse catene leggere e pesanti possono associarsi
per produrre almeno 10¹⁶ forme diverse. La variabilità si concentra nei domini
di legame per l'antigene delle immunoglobuline. Sebbene questa variabilità
innata non sia elevata come quella del recettore delle cellule T, le cellule B
possiedono un'altra via molto utile per raggiungere la specificità: la
mutazione somatica. Questa è raggiunta permettendo l'introduzione di errori durante
la trascrizione del DNA, caso che si verifica circa una volta ogni appaiamento
di mille coppie di basi. Questa frequenza è 100 volte maggiore della frequenza
di mutazione dell'RNA virale e suggerisce un interessante parallelo tra la
capacità di microevolvere del sistema immunitario e quella degli agenti
patogeni. Le cellule B contenenti errori che migliorano il legame con
l'antigene sono ricompensate da un aumento di stimolazione, crescita e
divisione cellulare. Le cellule che producono anticorpi che legano fortemente
particelle estranee proliferano, producendo una maggior quantità di anticorpi
che trovano così un bersaglio opportuno.
La seconda componente delle immunoglobuline è chiamata
frammento cristallizzabile (Fc, Fragment crystallizable). Per una data sotto
classe di anticorpi questa componente è costante e non è rivolta verso
l'antigene (fig. 4). Essa interagisce con il complemento
o con i propri recettori sulle cellule, determinando così l'effetto del legame
dell'antigene alla porzione Fab (Fragment antigen-binding, frammento che lega
l'antigene). l differenti tipi di immunoglobuline sono identificati in relazione
alla componente Fc e ciascuna ha un diverso ruolo. Così come la risposta
immunitaria si modifica nel tempo, anche le cellule B maturano e vanno incontro
allo switch isotipico. Lo stesso Fab codificato dai geni V H si unirà
successivamente con diversi prodotti dei geni CH, in modo da produrre IgM, IgG,
ecc. Le IgG costituiscono le più abbondanti immunoglobuline del siero e i
principali anticorpi presenti dopo l'immunizzazione con la maggior parte degli
antigeni. Non sono attivamente secrete nel tratto respiratorio ma travasano dal
circolo sanguigno quando la mucosa è infiammata. Le cellule B che producono IgG
di solito giacciono negli strati più profondi della mucosa. Le IgG sono molto
potenti nell'opsonizzare l'antigene che è inglobato dalle cellule polimorfonucleate
(PMN) e dai macrofagi. L'adesione è seguita dall'ingestione (fagocitosi) e
dalla fusione del fagosoma internalizzato con i lisosomi. Gli enzimi
proteolitici, i radicali dell' ossigeno e l' acidificazione del fagolisosoma
contribuiscono attivamente alla demolizione del contenuto del fagolisosoma
stesso.
l monomeri di IgM possono unirsi con le catene J che
legano le componenti secretrici sulla superficie basolaterale degli enterociti.
Questi anticorpi rilasciati complessano l'antigene che è quindi eliminato
dall'interno dell'organismo (esclusione immunitaria). Le IgM sono anticorpi
opsonizzanti assai potenti e sono presenti nel momento in cui le cellule B
incontrano l'antigene per la prima volta. Sono anche forti attivatori delle
componenti del complemento, che formano pori nelle cellule (lisi). La
componente inattiva del complemento, chiamata Clqrs (dove q, re s sono le tre
subunità del complesso Cl) può essere attivata dal legame a una singola
molecola IgM (o ad almeno due molecole IgG), che è andata incontro a un cambio
conformazionale dopo il legame con l'antigene. Questa a sua volta attiva Clq,
che attiva Clr, che attiva CIs (v. fig. 4c), iniziando la via classica di
attivazione del complemento (fig. 5).
Le IgA sono immunoglobuline specializzate presenti a
basse concentrazioni nel siero ma molto abbondanti nelle secrezioni mucose.
Sono attivamente trasportate attraverso le cellule epiteliali, dove si legano
alla componente secretrice, e quindi rilasciate nelle secrezioni (latte,
saliva, ecc.). La loro azione antivirale si esplica, per esempio, proteggendo
la mucosa intestinale dall' infezione da parte di poliovirus e il tratto
respiratorio dai comuni virus influenzali, inclusi il virus respiratorio
sinciziale e quello dell'influenza A. Le IgD sono presenti a livelli bassissimi
nel siero, ma si trovano sulla superficie di molte cellule B nelle fasi precoci
del differenziamento. Anche le IgE sono presenti a concentrazioni bassissime
nel siero, ma sono importanti perché la componente Fc si lega molto fortemente
ai mastociti, ai baso fili e agli eosinofili. Un antigene che si leghi alle IgE
fissate alla superficie di queste cellule provoca il rilascio di istamina,
bradichina, enzimi, leucotrieni, fattore attivante le piastrine, ecc. Si
ritiene che siano particolarmente importanti nella protezione contro infezioni
da parte di vermi, ma che inneschino anche reazioni allergiche, come, per
esempio, febbre da fieno o asma. Gli anticorpi contro i virus possono essere
neutralizzanti; in questo caso il legame dell'anticorpo blocca la capacità
infettiva del virus ostruendo fisicamente i siti critici per l'adesione e
l'ingresso del virus sulle proteine di superficie. In alternativa, l'anticorpo
può legare altri siti che non interferiscono con il legame del virus. Questo
anticorpo non neutralizzante può opsonizzare il virus direttamente o lasciarlo
legare dal complemento. Infine, alcuni virus sembrano infettare meglio in
presenza di un anticorpo, usando l'opsonizzazione per penetrare in cellule che
non sono capaci di distruggere il virus. Questi sono anticorpi che aumentano
l'infezione.
L'anticorpo, di qualsiasi tipo sia, è spesso inefficace
nel prevenire le infezioni virali delle superfici mucose; ciò dipende dal fatto
che esso non è specifico o è presente nel posto o nel momento sbagliati. Se le
cellule B non hanno ancora incontrato un determinato antigene virale, gli
anticorpi con la corretta specificità impiegano tempo per espandersi. Per
esempio, se il sistema immunitario non ha mai incontrato il ceppo di influenza
diffuso in un particolare anno, gli anticorpi contro altri ceppi sono
inefficaci contro le proteine di superficie del nuovo ceppo. Un caso recente è
stato l'esplosione dell'influenza aviaria H5Nl nel 1997 a Hong Kong. Alcune
altre specie di virus del raffreddore, per esempio i rhinovirus, sono in grado
di assumere un centinaio di rivestimenti diversi, costringendo il sistema
immunitario a trattare ogni infezione come se fosse nuova. Nel caso del virus
respiratorio sinciziale, inoltre, anticorpi presenti nel siero ai normali
livelli sono inefficaci contro la reinfezione per via nasale. Al contrario, IgA
localizzate nel naso sono efficaci, ma persistono solo per 9 mesi dopo
l'infezione. Purtroppo, dato che le esplosioni virali si succedono circa una volta
all'anno, questi anticorpi sono di poca utilità nella protezione contro le
ondate successive.
Processamento e presentazione degli antigeni virali
alle cellule T
Le cellule T riconoscono esclusivamente gli antigeni
che sono stati processati, cioè ridotti a livello di frammenti peptidici.
Questi frammenti sono presentati sulla superficie delle normali cellule legati
a specifici siti di proteine di superficie, chiamate MHC. Se le cellule sono
infettate da un virus, i nuovi peptidi endogeni, compresi alcuni peptidi di
origine virale, vengono campionati dall'MHC di classe I e portati sulla
superficie cellulare. Al contrario, alcune cellule specializzate che presentano
l'antigene (APC, Antigen Presenting Cells) prelevano proteine dal mezzo
extracellulare e le digeriscono in frammenti peptidici per la presentazione da
parte dell'MHC di classe II. Quando le cellule T incontrano questi nuovi
complessi MHC/peptide, vi si legano attraverso il recettore delle cellule T
(TCR, T Cell Receptor); il legame è stabilizzato da CD4, molecola attraverso la
quale le cellule T coadiuvanti legano la classe II, o da CD8, molecola
attraverso la quale i linfociti T citotossici (CTL, Cytotoxic T Lymphocytes)
legano la classe I. Singoli linfociti T maturi riconoscono un numero molto
limitato di antigeni. Il riconoscimento avviene attraverso l'appaiamento della
struttura della regione apicale del recettore delle cellule T, posto sulla
superficie della cellula T, con una struttura complementare dell'antigene
presentato da altre cellule ospiti.
Le più importanti cellule APC sono i macrofagi, le
cellule dendritiche e i linfociti B. La modalità con cui l' antigene entra
nelle APC è rilevante per determinare quale tipo di cellula T verrà stimolata
dall'antigene. Antigeni esogeni (solubili) vengono inglobati dalle APC, in modo
particolare quando sono opsonizzati dagli anticorpi o dal complemento, o quando
sono legati all' anticorpo sulla superficie delle cellule B. Dopo essere stato
intemalizzato, l'antigene viene demolito nei fagolisosomi. l peptidi derivati
dalla digestione si incontrano con le proteine dell'MHC di classe II neo
sintetizzate (o riciclate) che sono accompagnate, fino a questo punto, da una proteina
chiamata catena invariante (v. il saggio di G.J. Hämmerling e collaboratori,
nel II volume). La catena invariante si lega strettamente all'MHC di classe II
e si infila nel solco sull' apice della proteina di classe II, dove si è
formato il sito di legame per i peptidi prodotti dalla digestione
dell'antigene. Dopo che la catena invariante viene rimossa, il sito di legame
per il peptide cattura un qualsiasi peptide disponibile che si adatti al solco.
Se non sono disponibili peptidi estranei, l'MHC II cattura peptidi normali
della cellula ospite prodotti dal naturale riciclo delle proteine in questo
comparto della cellula. Ogni proteina MHC II può potenzialmente trattenere
diverse migliaia di peptidi differenti. Sebbene sembri un numero enorme, appare
piccolo se confrontato con il numero potenziale di peptidi presenti in ogni
cellula. Questi peptidi sono di solito lunghi 15 ÷18 amminoacidi e sporgono per
un breve tratto a una delle estremità del solco dell'MHC II. Si pensa che circa
100 o 200 proteine MHC debbano essere occupate dal peptide appropriato perché
avvenga il riconoscimento. Questa quota rappresenta 0,1 ÷ 10% delle proteine
MHC presenti sulla superficie di una APC.
La via di presentazione degli antigeni endogeni mostra
alcune importanti differenze rispetto alla presentazione di antigeni esogeni.
Gli antigeni endogeni sono sintetizzati dentro la cellula e quindi demoliti in
peptidi di 8 ÷ 10 amminoacidi. Questi peptidi più corti incontrano la proteina
MHC di classe I mentre essa viene prodotta nel reticolo endoplasmatico rugoso e
l'associazione del peptide con l'MHC l aiuta a stabilizzare il legame con la
β2-microglobulina. Il complesso trimolecolare si muove attraverso l' apparato
del Golgi, dove le proteine vengono glicosilate prima di essere esposte in
superficie. l peptidi si adattano al solco all'apice dell'MHC l in modo quasi
analogo ai peptidi che si adattano all'MHC 11, tranne per il fatto che il solco
dell'MHC l è chiuso a entrambe le estremità. Ciò implica che ciascun MHC l ha
una scelta più limitata di peptidi che vi si adattano in modo adeguato: per
questa ragione, vengono presentate solo poche centinaia di peptidi diversi
entro il solco degli MHC l sulla superficie di cellule caratteristiche. Questo
repertorio limitato è compensato, in parte, da una gamma più vasta di cellule
che esprimono l'MHC I. La maggior parte delle cellule nucleate porta l'MHC l,
incluse tutte le cellule dell' epitelio delle mucose, le cellule che presentano
l'antigene, i fibroblasti e l'endotelio dei vasi. Al contrario, l'MHC II è
presente in quantità importanti solo su cellule APC 'professioniste'. Le due
proteine MHC funzionano perciò in due modi fondamentalmente diversi. L 'MHC di
classe I mostra quali proteine la cellula sta sintetizzando e invita i linfociti
T citotossici a uccidere la cellula che mostra complessi MHC/peptide anomali.
L'MHC di classe II è invece presente solamente sulle cellule che sono
specializzate nel 'diffondere notizie', che campionano le proteine presenti nel
mezzo extracellulare e innescano una risposta immunitaria coordinata mediata
dalle cellule T helper.
La capacità delle cellule T di discriminare tra il self
e il non self, cioè tra ciò che è proprio dell' organismo e ciò che è estraneo,
si sviluppa nel timo. Dopo aver lasciato il midollo osseo, le cellule destinate
a diventare cellule T migrano nel timo; qui vanno incontro a una serie di
cambiamenti nel passaggio dallo strato corticale a quello midollare. Il timo è
effettivamente una 'stazione di esame', progettata per selezionare le cellule T
con bassa reattività verso gli MHC, che presentano antigeni del self ma sono
peraltro capaci di riconoscere peptidi estranei presentati da MHC propri.
Appena dopo il loro arrivo, le cellule migrate dal midollo osseo mostrano sulla
propria superficie sia CD4 sia CD8. Quando entrambe queste proteine sono
presenti con un complesso TCR, la forza di interazione con le cellule che
presentano l'antigene nello stroma del timo determina il destino della cellula
T in via di sviluppo. Quelle che si legano troppo fortemente o troppo
debolmente vengono eliminate o entrano in uno stato di inattività
semipermanente, denominata anergia. Questo processo di selezione timica è molto
rigoroso e la maggior parte delle cellule T in sviluppo muore nel timo per
apoptosi. Le cellule T selezionate per essere di potenziale utilità futura per
l'ospite si moltiplicano ed escono dal timo; circolano poi attraverso i
linfonodi, la milza, il fegato, l'intestino, la cute e altre regioni, cercando
nuovi complessi MHC/peptide sulla superficie delle cellule ospiti. Una volta in
periferia, le opportunità di ulteriori cambiamenti nelle proteine di superficie
delle cellule T sono estremamente scarse. Queste cellule T hanno un destino ben
stabilito e difficilmente mutabile: infatti, hanno perso la capacità di
esprimere altri corecettori e non possono modificare il loro TCR. Esse restano
quiescenti fino a quando vengono in contatto con una cellula che possiede le
caratteristiche per il cui riconoscimento sono programmate e solo allora si attivano.
Il processo attraverso il quale le cellule T raggiungono le cellule che
presentano l' antigene non è soltanto casuale, sebbene la frequenza giochi una
parte quando vi sono solo pochissimi linfociti con un TCR che si adatta
intimamente con qualsiasi complesso MHC/peptide. Nelle zone in cui avviene
l'infezione, l'infiammazione provoca un aumento del flusso sanguigno, un
travaso più attivo di linfociti nei tessuti e un'attivazione delle APC, che
aumenta la quantità di proteine MHC sulla superficie. Quando una cellula che
presenta l'antigene incontra il TCR complementare, la cellula T è portata in
uno stato di attivazione metabolica, che culmina con la divisione cellulare. Il
processo di sviluppo dei linfociti è perciò un equilibrio finemente controllato
tra crescita, inattività, attività e morte. Tutte queste caratteristiche sono
essenziali per un corretto funzionamento del sistema immunitario. Lo sviluppo
dei linfociti corrisponde, in un certo senso, allo sviluppo e alla capacità
adattativa degli organismi patogeni, contro cui il sistema immunitario si è
evoluto. La diversità potenziale dei TCR è immensa. Si è calcolato che i geni
per i TCR sono capaci di unirsi in modi diversi per generare circa 10¹⁶
specificità differenti, assicurando che uno o più TCR saranno quasi
perfettamente adatti a ogni proteina estranea. Molte di queste possibilità
saranno rigettate dal timo perché avranno una interazione troppo forte o troppo
debole con le normali cellule ospiti. In ciascuno di noi vi è stata una corsa
evolutiva effettuata dai nostri linfociti, già prima della nascita, attraverso
la morte per apoptosi di circa il 95% delle cellule che hanno iniziato il
programma differenziativo. Durante tutta la vita, i linfociti continuano a
essere selezionati, resi tolleranti o eliminati. Una quota relativamente esigua
di cellule T sopravvive negli anziani e questo spiega forse in parte le
capacità limitate del sistema immunitario di rispondere alle infezioni in età
avanzata.
Cellule T helper
La presenza di CD4 sulla superficie di una cellula T
provoca un grande rafforzamento del legame del recettore delle cellule T
all'MHC di classe II che porta il peptide 'giusto'. Quando le cellule vergini
uscenti dal timo entrano in contatto con una cellula che presenta l'antigene,
iniziano a proliferare e a produrre segnali chimici sotto forma di citochine.
Esempi importanti di citochine prodotte da queste cellule T helper sono IL-l,
lL-3, lL-4, IL-5, IL-10 e interferone γ. Queste sostanze chimiche segnalano ad
altre cellule di prepararsi ad attaccare l'agente patogeno invasore. lL-3 è un
fattore chiave che provoca la crescita e la divisione delle cellule staminali,
producendo più cellule B e T. lL-2 ha un ruolo chiave nell'attivazione delle
cellule T, mentre lL-4 e lL-6 attivano le cellule B. lL-3, in combinazione con
GM-CSF (Growth Macrophage Colony Stimulating Factor, fattore che stimola la
crescita della colonia di macrofagi), è un potente promotore della crescita e
della proliferazione delle cellule fagocitiche, compresi eosinofili, polimorfonucleati,
baso fili e macrofagi. Le cellule T helper giocano un ruolo centrale
nell'identificare in modo preciso l'agente patogeno invasivo e nell'accendere
il sistema d'allarme dell'ospite che coinvolge altri giocatori-chiave della
difesa immunitaria.
Le cellule T helper possono essere classificate in tipi
diversi, che producono combinazioni caratteristiche di citochine: alcune
combinazioni dirigono le risposte immunitarie, mentre altre attivano la
produzione di anticorpi da parte delle cellule B (Fiorentino et al., 1989;
Mosmann e Sad, 1996; Street et al., 1990).
Alcuni antigeni e diverse modalità di immunizzazione
inducono in modo differenziale due principali sotto gruppi funzionali, THl e
TH2 (fig. 6). Le cellule THl producono cito chine
di tipo l, che includono TNF-α (Tumor Necrosis Factor α, fattore di necrosi
tumorale α) e lFN-γ; queste hanno un potente effetto antivirale. Le cellule TH2
producono cito chine di tipo 2 che comprendono lL-4, lL-5 e IL-10 e sono
predominanti nelle risposte antielmintiche e allergiche. Popolazioni di cellule
che producono citochine di tipo l e 2 sono mutualmente inibitorie. Sebbene sia
stata osservata produzione polarizzata di citochine in cloni di cellule T
provenienti da parecchie specie (sia di primati che di roditori), il grado di
polarizzazione nelle cellule T naturali si sta chiarendo solo ora.
Cellule T citotossiche
La via di processamento endogena porta dapprima
all'attivazione delle cellule T con CD8, che funzionano per lo più come cellule
T citotossiche. Di nuovo, il processo di coinvolgimento della cellula T
'giusta' consiste essenzialmente nel trovare un TCR con la struttura adeguata
per adattarsi al complesso MHC/peptide. Una volta avvenuto il contatto, la
presenza di CD8 sulla superficie della cellula T blocca la cellula T stessa
sull' APC che porta il peptide appropriato. La risposta delle cellule T CD8+
(cellule T che esprimono CD8) è di nuovo l'attivazione (fig. 7); sebbene tali cellule producano
anche citochine, soprattutto di tipo l, l'effetto principale dell'attivazione è
che le cellule diventano citotossiche. l CTL attivati riorientano il cito
scheletro e l'apparato secretore verso la cellula bersaglio e rilasciano
granuli contenenti perforina e granzimi nello spazio formatosi tra le due
cellule. La perforina forma polimeri in presenza di Ca²+ e si inserisce nel
doppio strato lipidico della membrana della cellula bersaglio, formando pori
delimitati da polimeri di perforina. Il conseguente ingresso di acqua e ioni
provoca lo scoppio della cellula ospite. La cellula allora frammenta il suo DNA
e il suo RNA e muore. A questo punto, la cellula T si stacca dalla cellula
morente, si divide e si sposta per cercare altre cellule da contattare e
uccidere. È importante che le cellule T citotossiche siano altamente selettive
nella scelta dell'antigene; l'uccisione incontrollata di cellule erroneamente
riconosciute come infettate sarebbe infatti disastrosa.
Un altro meccanismo citotossico, il principale
utilizzato dai CTL che non possiedono granuli, consiste nell' espressione del
ligando per Fas (FasL) indotta dall'attivazione. FasL si lega al suo recettore
Fas presente sulla membrana della cellula bersaglio, portando bersaglio e CTL
molto vicini. Il legame innesca l'apoptosi, che conduce alla frammentazione del
DNA e alla morte cellulare. l CTL si staccano dal bersaglio dopo aver scagliato
il colpo mortale e riescono così a uccidere più cellule in rapida successione.
Esempi di risposte immunitarie a particolari agenti
patogeni
Virus respiratorio sinciziale
Le infezioni polmonari sono la causa dominante di
morbilità e mortalità infantile. Il virus respiratorio sinciziale (RSV,
Respiratory Syncytial Virus) è il principale agente patogeno dell'apparato
respiratorio infantile a livello mondiale e rappresenta una sfida irrisolta per
lo sviluppo di un vaccino (Heilman, 1990). Dal punto di vista clinico,
l'infezione è caratterizzata da sintomi e segni di restringimento bronchiale e molti
bambini che si sono ristabiliti dalla bronchiolite sono stati successivamente
diagnosticati asmatici. Le difese immunitarie contro RSV sono interessanti per
molti aspetti. Innanzi tutto, si verificano nuove infezioni durante la vita
malgrado risposte immunitarie apparentemente forti alla prima infezione.
Inoltre, la vaccinazione può aumentare la gravità della malattia. Infine, vi
sono legami apparenti tra le infezioni infantili e il successivo sviluppo
dell'asma. RSV appartiene al genere Pneumovirus della famiglia Paramyxoviridae
ed è simile al virus Marburg, al virus del morbillo, al virus del cimurro
canino e ai virus della parotite e della parainfluenza. Il microscopio
elettronico mostra virioni di forma irregolare e spesso aggregati, con un
involucro lipidico che porta le glicoproteine di superficie G, F e SH (v.
oltre). Il nucleocapside contiene un genoma a RNA a singolo filamento negativo
(vengono designati a filamento negativo quei virus che trascrivono l'RNA
messaggero necessario per la sintesi delle proteine a partire dall'RNA genomico
e replicano quest'ultimo tramite un RNA complementare di senso positivo), di
massa molecolare 5 ∙ 10⁶ kDa, non segmentato. Sono presenti 10 geni con 12
potenziali prodotti. La trascrizione procede sequenzialmente da 3' a 5'; i
primi geni trascritti sono lc e lb, che codificano proteine non strutturali con
funzione sconosciuta. Viene poi trascritta la nucleoproteina (N), che è
relativamente ben conservata tra ceppi isolati naturali. La fosfoproteina (P) e
la piccola proteina idrofobica (SH) sono trascritte di seguito, seguite dalla
proteina di adesione (G) e dalla proteina di fusione (F). Queste ultime due
proteine sono le principali glicoproteine superficiali, contro cui sono diretti
gli anticorpi neutralizzanti, e mostrano la maggiore variabilità naturale tra
diverse forme isolate naturali di RSV. Infine vi sono M2, seconda proteina di
matrice, di 22 kDa, e la proteina large (L) che è la RNA-polimerasi. Si pensa
che entrambe queste proteine siano relativamente ben conservate. Studi
sull'uomo e su animali modello hanno mostrato il doppio ruolo delle cellule T
antivirali nell'eliminare il virus e nel provocare malattie più acute. Questo
paradosso immunopatologico è ora compreso in modo più chiaro sia per la
malattia provocata da RSV sia per quelle provocate da qualsiasi altra comune
infezione umana.
La prima prova che l'immunità specifica potesse essere
pericolosa si è avuta negli anni Sessanta, quando dei bambini furono vaccinati
con RSV inattivato dalla formalina. l bambini riceventi il vaccino svilupparono
forti risposte sierologiche, ma non furono protetti dall'infezione. Molti di
questi individui, che vennero in seguito in contatto con RSV, svilupparono una
grave affezione del tratto respiratorio inferiore e alcuni ne morirono. L'esame
post mortem dei soggetti vaccinati col virus inattivato mostrava
un'infiltrazione di eosinofili. Si sono a lungo studiate le cause
dell'affezione aggravata dal vaccino inattivato, ma purtroppo non si è ancora
prodotto un vaccino sicuro ed efficace. Recentemente sono state riesaminate le
risposte immunitarie a RSV (Openshaw, 1995). L'immunità umana naturale a
seguito di un'infezione ha durata breve e durante la vita si possono verificare
nuove infezioni. Dopo i deludenti risultati dei primi tentativi con RSV
inattivato dalla formalina, si sono proposti vaccini alternativi nel tentativo
di indurre risposte protettive nella mucosa respiratoria. Tra questi sono stati
testati ceppi attenuati di RSV, vaccini peptidici o derivati costituiti da singole
subunità incorporati in sostanze adiuvanti in grado di potenziare le risposte
delle mucose, adenovirus ricombinanti o virus vaccini che esprimono le proteine
F o G, o entrambe, di RSV. A tutt'oggi, l'efficacia di questi protocolli di
vaccinazione non è ancora stata determinata nell'uomo. Nel modello murino per
l'affezione da RSV, la maggior parte dei linfociti recuperati durante i primi 5
giorni dall'infezione primaria è di fenotipo CD4- e CD8- e presenta le
caratteristiche delle cellule NK, la cui attività mostra un picco circa nello
stesso momento. Durante l'eliminazione del virus dai polmoni (6 - 9 giorni), il
principale sottogruppo di linfociti è di tipo CD8+, sebbene si trovino anche
cellule CD4+. In questo modello, l'induzione di forme distinte di
immunopatologie può essere spiegata dalla produzione di citochine di tipo 2
dalle cellule T, che riconoscono la principale glicoproteina di superficie G, e
di citochine di tipo l da parte di cellule T specifiche per la proteina di
fusione F e per le proteine della matrice M2. La prova più convincente che
l'aggravarsi della malattia sia causato dalle cellule T viene da studi sul
trasferimento passivo dell'immunità. Topi infettati solo con RSV mostrano una
lieve infermità, ma la gravità della malattia aumenta drammaticamente con
l'iniezione di cellule T che riconoscono RSV. Le cellule T H l o i CTL CD8
possono provocare emorragie polmonari (che ricordano il polmone da shock),
mentre le cellule CD4, producendo citochine di tipo 2, provocano eosinofilia
nei polmoni (Alwan et al., 1994). Al contrario, il trasferimento di anticorpi
non aggrava mai la malattia in vivo e può talvolta proteggere dall'infezione.
Diversi studi hanno suggerito una relazione tra la
bronchiolite da RSV e l'asma. N. Sigurs e collaboratori (1995) hanno realizzato
uno studio prospettivo di gruppo sulla bronchiolite da RSV infantile e hanno
scoperto che essa è uno dei maggiori fattori di rischio per lo sviluppo di asma
e malattie atopiche. Da questi studi non emerge una chiara indicazione che la
bronchiolite operi come fattore di rischio indipendente, ma sembra sia un
fattore aggiuntivo che agisca indipendentemente dalla storia familiare dell'
atopia. Il tipo di risposta immunitaria che si verifica nel contatto iniziale
con RSV dipende da parecchi fattori; tra questi le influenze genetiche, lo
stato immunologico, la presenza di risposte immunitarie in atto contro altri
antigeni e la presenza di memoria di antigeni simili.
HIV e AIDS
L'infezione provocata dal virus dell' immunodeficienza
umana (HlV, Human lmmunodefieieney Virus) non è in genere eliminata dal sistema
immunitario. Dopo esser penetrato nell'organismo, HlV costituisce rapidamente
delle riserve di virus al di fuori della portata dei meccanismi immunitari di
eliminazione. Queste riserve sono:
1) infezione virale latente di cellule T CD4+
quiescenti; nell'organismo si genereranno nuovi virus dopo l'attivazione delle
cellule T.
2) Virioni ricoperti dagli anticorpi e dal complemento
intrappolati nella rete delle cellule dendritiche follicolari nei linfonodi;
questi virioni non sono neutralizzati ma, al contrario, sono altamente
infettivi e sono concentrati nei siti obbligati di ricircolazione delle cellule
T CD4+.
3) Varianti che hanno perso gli epitopi antigenici e
sfuggono al controllo immunitario. Inoltre, l'instabilità del genoma virale è
alta e varianti del virus si sviluppano per tutta la durata dell'infezione in
diverse regioni anatomiche, costituendo un bersaglio in continuo cambiamento
per meccanismi di controllo effettori. Ne consegue che molti processi
immunitari vengono attivati durante l'infezione, ma non riescono a estirpare il
virus.
Dopo l'ingresso di HlV, l'infezione si diffonde
rapidamente alle cellule che esprimono CD4 e i recettori per le chemochine CCR5
e CXCR4 (Hill et al., 1997). Le risposte cellulari e anticorpali seguono la
prima massiva ondata di viremia che scaturisce dalla replicazione virale. La
quantità di cellule T CD4+ circolanti crolla contemporaneamente al picco della
viremia; questo è in relazione al fatto che i CD4+ sono gli ospiti principali
per la replicazione di HlV. Il picco della risposta dei CTL è seguito da un
crollo nella viremia e dal ritorno a livelli normali delle cellule CD4+; ciò
suggerisce che almeno temporaneamente i CTL sono capaci di controllare HIV. La
risposta degli anticorpi neutralizzanti compare più tardi e arriva lentamente a
plateau. Questa fase acuta dell'infezione dura circa 3 mesi ed è seguita da una
fase cronica, durante la quale le risposte dei CTL restano molto attive, il livello
degli anticorpi è alto, quello dei virus circolanti è basso e le cellule CD4+
scendono lentamente. La fase cronica può durare parecchi anni ed è
caratterizzata da un veloce e costante riciclo dei virus, che fa diminuire
progressivamente il numero delle cellule CD4+ circolanti (Ho et al., 1995). Con
il tempo si verifica, nella gran parte degli individui infettati, una
catastrofica transizione alla sindrome da immunodeficienza acquisita (AIDS,
Acquired lmmunodeficiency Syndrome): il numero delle cellule CD4+ circolanti
crolla sotto la soglia critica (<200/μl), i CTL e gli anticorpi scompaiono e
l'ospite alla fine soccombe a causa di infezioni opportunistiche o di tumori
correlati all'AIDS.
Individui con varianti genetiche delle molecole dei
corecettori virali mostrano resistenza all'infezione e alla progressione verso
l'AIDS (Liu et al., 1996). Questo non è dovuto a meccanismi immunitari
effettori, anche se i recettori delle chemochine coinvolti hanno un ruolo nelle
risposte immunitarie, ma riflette semplicemente il debole legame alle cellule.
Altri fattori coinvolti nella progressione lenta verso l'AIDS potrebbero essere
risposte protettive finora non identificate, inoculi scarsamente infettivi o
potrebbero semplicemente rappresentare l'estremo limite di una gamma di
velocità di progressione.
Cellule CD4+. - Il decremento delle cellule CD4+ è probabilmente
dovuto a una combinazione tra gli effetti citopatici virali e l'uccisione da
parte di HIV dei CTL che riconoscono i peptidi di HIV espressi in associazione
a molecole MHC di classe I sulla membrana delle cellule CD4+ infettate. Anche
prima che un calo nel numero delle cellule CD4+ divenga evidente, si riscontra
una riduzione delle risposte immunitarie ad alcuni altri stimoli. Con il tempo,
si perde la capacità di rispondere agli alloantigeni e alla fine anche quella
ai mitogeni. Si sono osservate la perdita precoce delle risposte di tipo THl
(produzione di IFN-γ e IL-2) e la ritenzione preferenziale delle risposte di
tipo T H2 (produzione di IL-4 e IL-10) durante il progredire verso l'AIDS. Ciò
può essere dovuto al fatto che le cellule T H l sono più suscettibili
all'uccisione da parte di HIV delle cellule TH2, o all'induzione positiva delle
cellule TH2 mediata da HIV. Si è osservato anche il coinvolgimento di cellule
CD4+ citotossiche, che riconoscono i peptidi derivati da gpl20 presentati dalle
molecole MHC di classe II, ma non è chiara la loro importanza.
Cellule CD8+. - I livelli di CTL circolanti specifici per HIV sono
estremamente alti (circa l'1% dei linfociti) durante tutta la fase cronica
(Moss et al., 1995), al punto che è possibile saggiare direttamente la loro
attività in saggi citotossici ex vivo. Di solito sono presenti più cloni di CTL
che riconoscono differenti antigeni virali e si sono osservate dinamiche
complesse di espansione clonale delle cellule CDS+ e del carico di virus
mutanti (Nowak et al., 1995). Oltre all'attività citotossica, i CTL possono
rilasciare IFN-γ (effetto antivirale diretto) e chemochine come RANTES, MIP-l α
e MIP-lβ, che possono competere con HIV per i corecettori usati dal virus per
penetrare nelle cellule CD4+ e infettarle. Questa competizione impedisce
l'ingresso di virus libero in nuove cellule ospiti, anche se questo non è un
effettore diretto e non è in grado di eliminare l'infezione.
Anticorpi. - Le
risposte anticorpali si sviluppano più lentamente di quelle dei CTL, forse
perché necessitano dell'aiuto da parte delle cellule CD4+. I principali
anticorpi neutralizzanti sono diretti contro la glicoproteina (gp) virale di
rivestimento gp120, in particolare contro un'ansa molto variabile chiamata V3.
Il rapporto tra mutazioni codificanti e non codificanti dell'ansa V3 suggerisce
che essa sia sotto una pressione selettiva, forse una combinazione tra la
neutralizzazione da parte degli anticorpi e la disponibilità di molti
corecettori alternativi. Non è chiaro quale sia l'esatto meccanismo di
neutralizzazione. Non tutti gli anticorpi antiansa V3 sono neutralizzanti; gli
anticorpi neutralizzanti che riconoscono l'ansa V3 non impediscono il legame a
CD4, ma potrebbero bloccare la fase di ingresso del virus dopo il legame. Un
altro meccanismo possibile è l'inibizione del taglio della glicoproteina
dell'involucro, un passaggio richiesto per l'infettività mediato da una proteasi
che trasforma il precursore gpl60 env in gpl20 e gp41. Altri anticorpi
neutralizzanti riconoscono il dominio di legame per CD4 su gpl20 e permettono
la neutralizzazione crociata di ceppi virali con diverse anse V3. Anche altre
regioni di gpl20 sono bersaglio degli anticorpi neutralizzanti che possono
funzionare alterando la conformazione del virione o interferendo con il legame
ai corecettori. In sistemi sperimentali che utilizzano gli scimpanzé, alti
livelli di anticorpi neutralizzanti che riconoscono la glicoproteina
dell'involucro possono proteggere contro l'infezione da HIV, ma tali livelli
non possono essere sostenuti per lungo tempo. Inoltre, variazioni
nell'involucro virale possono impedire tale protezione. Nelle prove cliniche di
vaccini basati sull'involucro virale, gli anticorpi prodotti non riescono a
neutralizzare i ceppi isolati primari, mostrando le limitazioni della
neutralizzazione ceppo-specifica.
ADCC. - È
stata anche descritta una forma di citotossicità cellulare (ADCC, Antibody
Directed Cellular Cytitoxicity) mediata dagli anticorpi che riconoscono le
proteine dell'involucro gpl20 e gp41 e che prevede il coinvolgimento delle
cellule NK. Il ruolo di questo meccanismo nel controllo della replicazione di
HIV dipende dall' attività delle cellule NK, poiché gli anticorpi sono presenti
ad alta concentrazione durante gran parte della fase cronica. Questa attività
può essere meno utile dei meccanismi di citotossicità mediati dai CTL, o
addirittura dannosa, perché potrebbe causare la lisi delle cellule T CD4+ non
infettate che hanno legato la gpl20 rilasciata, o delle cellule B che esprimono
anticorpi di membrana che riconoscono gp120.
Anticorpi potenzianti. - I complessi circolanti tra HIV e anticorpi sono
infettanti e non vengono distrutti dai macrofagi. Infatti, gli anticorpi
possono aumentare l'infettività di HIV. Questo si verifica per legame diretto
dei complessi immunitari o alle cellule che esprimono il recettore per Fc,
soprattutto CDl6 sulle cellule NK e il recettore FcRI sui monociti, oppure alle
cellule che esprimono CR2 per il potenziamento mediato dal complemento. Tale
potenziamento si dimostra con sieri neutralizzanti che sono stati diluiti oltre
la concentrazione richiesta per la neutralizzazione. Così i livelli decrescenti
di anticorpi nel decorso della malattia possono facilitare il progredire verso
l' AIDS. Inoltre, anticorpi che neutralizzano un ceppo di HIV possono
potenziarne un altro. Questo effetto può contribuire alla selezione di varianti
resistenti ed evidenzia un possibile problema per le vaccinazioni indirizzate
all'induzione dell'immunità mediata dagli anticorpi.
Cellule NK. -
L'attività delle cellule NK sembra diminuire con il progredire verso l'AIDS, ma
l'aggiunta di IL-2 può far recuperare l'attività nei saggi in vitro di funzione
delle NK stesse. Anche se un sistema immunitario senza aiuti sembra essere
tutt'al più capace solo di rallentare la progressione della malattia, potrebbe
ancora essere possibile contrattaccare la variabilità e il vantaggio cinetico
di HIV mediante l'immunizzazione preventiva. Parecchi lavori hanno descritto
risposte cellulari specifiche all'RIV in individui che, pur essendo stati
esposti a tale virus, rimangono sieronegativi, anche se è stato difficile
mostrare l'effettiva eliminazione di un inoculo virulento di RIV. Si è
raggiunta protezione contro le infezioni da HIV in scimpanzé vaccinati e contro
le infezioni con il virus correlato dell'immunodeficienza di scimmia nei
macachi (Daniel et al, 1992). Il recente successo della terapia antivirale
combinata nella diminuzione della carica virale negli individui infetti
suggerisce che un trattamento antivirale precoce ed aggressivo dopo l'infezione
potrebbe prevenire l'esaurimento delle forze immunitarie, permettere lo
sviluppo di un'immunità protettiva solida e infine controllare la restante
replicazione virale (Autran et al., 1997).
Infezioni da Herpes simplex
Il virus Herpes simplex (HSV) ha un genoma a DNA
lineare a doppio filamento che codifica 70 o più proteine virali. Il genoma è
contenuto in un core (capside) circondato da un materiale amorfo, il tegumento,
ricoperto a sua volta da un involucro lipidico costellato di glicoproteine. Vi
sono due tipi di questo virus: HSV-l e HSV-2. Il secondo è di solito associato
con infezioni genitali.
HSV provoca una vasta gamma di malattie, dalle più
comuni, come herpes labiale, gengivostomatite, herpes genitale,
cheratocongiuntivite, alle infezioni viscerali di ospiti immunocompromessi e
alle rare encefaliti erpetiche. Negli USA, il 60 ÷ 80% della popolazione entro
i 50 anni è stato infettato da HVS-l e il 20% da HSV-2. HSV infetta per
contatto attraverso le mucose o le abrasioni cutanee. L'infezione è seguita
dalla replicazione nelle cellule epiteliali del sito di ingresso. A questo
punto HSV entra nelle terminazioni periferiche dei neuroni sensitivi e viene
trasportato dal flusso assonale retrogrado alle radici dei gangli dorsali, dove
va incontro a una replicazione limitata e transeunte. L'infezione neurale è di
solito controllata, anche se si verificano infezioni letali a carico del
sistema nervoso centrale in individui immunocompromessi, e il virus resta
latente, cioè il genoma si mantiene in forma episomica, non vengono sintetizzate
proteine virali e l'espressione genica virale è limitata ai trascritti
associati alla fase latente. Fattori di stress, come l'esposizione ai raggi
ultravioletti, stress emotivi, febbre, altre infezioni (per esempio, polmonite
pneumococcica), immunosoppressione, taglio di un nervo periferico, possono
essere seguiti dalla ricomparsa del virus nei siti innervati dagli stessi
neuroni infetti. Qui il virus entra nel ciclo litico replicativo a livello
delle cellule epiteliali, producendo vescicole e ulcere. In questo stadio si
verifica la trasmissione ad altri individui. Sono frequenti variazioni
individuali della frequenza di riattivazione, da una volta ogni pochi anni a 10
÷ 20 volte all'anno. La frequenza di infezioni ricorrenti (riattivate) potrebbe
dipendere dalla carica virale iniziale così come da fattori dell'ospite,
compresa la risposta immunitaria. HSV potrebbe diffondersi dal sito mucocutaneo
di ingresso attraverso i neuroni sensitivi per causare un'encefalite letale, ma
la maggior parte degli ospiti oppone resistenza alle affezioni diffuse
provocate da HSV. Le infezioni in ospiti che non sono mai stati esposti al
virus precedentemente provocano risposte infiammatorie locali nei siti
mucocutanei di replicazione. Per il precoce contenimento della diffusione del
virus nell'epitelio sono richieste la produzione locale di interferone di tipo
I (IFN-α e IFN-β) e le cellule NK. Oltre a dirigere gli effetti antivirali, gli
interferoni aumentano l'espressione dell'MHC di classe I e la presentazione
antigenica nei cheratinociti, fornendo un migliore bersaglio per i CTL.
Tuttavia, questi meccanismi effettori iniziali limitano ma non prevengono né
l'infezione dei neuroni né lo stabilirsi della latenza. L' eliminazione
dell'infezione locale richiede l'intervento dei linfociti T.
Esperimenti in modelli murini di infezione da HSV hanno
rivelato la complessità delle interazioni tra fattori genetici nell'ospite e
risposte delle cellule CD4+ e CD8+. In generale, l'eliminazione di HSV dal sito
primario di infezione dipende dalle cellule T CD4+, mentre le cellule T CD8+ ne
limitano la diffusione nei neuroni. Lo sviluppo di cellule T CD8+ specifiche
per HSV dipende dalle cellule CD4+ in alcuni, ma non in tutti, ceppi di topi.
La deplezione selettiva delle cellule CD4+ o CD8+ nell'animale produce effetti
limitati sull'eliminazione di HSV dalle lesioni primarie, mentre il trattamento
dei topi con anticorpi che neutralizzano IFN-γ, secreto da entrambi i sottotipi
di cellule T, prolunga la durata dell'infezione locale.
Le cellule T CD8+ si infiltrano nei gangli neuronali
con HSVe sembrano essenziali per controllare l'infezione neuronale, poiché il
trattamento con anticorpi anti-CD8 provoca una maggiore distruzione dei neuroni
(Simmons e T scharke , 1992). Il meccanismo di azione delle cellule T CD8+
specifiche per HSV nei gangli infetti è di tipo non litico: in realtà esse
aumentano il numero di neuroni che contengono il virus. Questo può essere
dovuto al fatto che l'espressione delle molecole MHC di classe I, normalmente
assenti nel tessuto nervoso, è bloccata a livello postrascrizionale nei
neuroni. Tuttavia, le cellule CD8+ riconoscono i peptidi virali prodotti in una
cellula infettata e presentati dalle molecole MHC di classe I di quella
cellula. L'infezione incrementa l'espressione della classe I nei gangli, ma
solo nelle cellule 'satellite' e in quelle di Schwann (Pere ira et al., 1994).
Diversamente dai neuroni, le cellule 'satellite' non sono permissive per la
replicazione di HSV, ma possono essere sede di un'infezione abortiva che
permette la presentazione dei prodotti virali sintetizzati nella fase iniziale
del ciclo replicativo. La sequenza di eventi dovrebbe quindi essere la
seguente: l'infezione litica dei neuroni rilascia il virus che infetta le
cellule 'satellite' senza successo, ma stimola da parte loro l'espressione
delle molecole MHC di classe I. Esse presentano i peptidi virali alle cellule
CDS+ infiltranti attratte dalle chemochine rilasciate durante le fasi iniziali
dell'infezione. Le cellule T CDS+ attivate rilasciano poi IFN-γ, che agisce sui
neuroni neo infettati e modula il ciclo replicativo del virus dalla fase litica
alla latenza. L' espressione dell'MHC di classe I cessa durante la fase
latente, quando si verifica solo la minima trascrizione dei geni virali e
diminuisce lo stato di attivazione immunitaria.
La latenza può durare per mesi o anni, finché stimoli
idonei provocano la riattivazione del virus. A questo punto la sintesi delle
proteine virali riprende e il virus infettivo ritorna indietro lungo le fibre
sensitive periferiche, fino ai siti mucocutanei innervati da tali fibre. Qui la
replicazione virale causa effetti citopatici evidenti a un esame clinico, cioè
vescicole trasparenti e ulcere. In circa il 50% dei casi di riattivazione,
tuttavia, la diffusione del virus è asintomatica, favorendo così la sua
trasmissione. In questa fase vengono innescati vari meccanismi dell'immunità
acquisita: le riattivazioni hanno durata più breve delle lesioni primarie. Gli
anticorpi neutralizzanti contro le glicoproteine dell'involucro (soprattutto gD
e gB) limitano la diffusione dell'infezione mediata da virus libero. HSV ha
evoluto misure contro gli anticorpi e il complemento. Le glicoproteine
dell'involucro gE e gl sono in effetti recettori dell'Fc che legano IgG
monomeriche, mentre gE da sola può legare aggregati di IgG. Tutto ciò sembra
proteggere i virioni dalla neutralizzazione. In modo simile, gC ha una parziale
omologia con il recettore del complemento CRI e lega C3b. Questo può aiutare
HSV ad attaccarsi a certi tipi cellulari.
Cellule mononucleate, compresi i linfociti CD4+ e CDS+,
si infiltrano nelle lesioni dovute all'infezione ricorrente. Le cellule CD4+
predominano nelle lesioni precoci. I linfociti T CD4+ e CDS+ specifici per HSV
sono presenti a bassa frequenza nelle PBMC (Peripheral Blood Mononuclear Cells,
cellule mononucleate del sangue periferico). Il clonaggio di cellule T isolate
da pazienti affetti da lesioni ricorrenti ha mostrato che i CTL riconoscono
parecchi antigeni diversi, derivati da proteine virali sia strutturali sia non
strutturali. In questi studi si sono usate linee di cellule B linfoblastoidi
infettate da HSV come cellule bersaglio. Quando sono stati utilizzati come
bersaglio i fibroblasti, la lisi mediata dai CTL è apparsa meno efficace.
Questo ha portato alla scoperta che HSV contiene un gene, denominato rx47, che
codifica il prodotto ICP4 7, la cui espressione può far trattenere l'MHC di
classe I nella cellula mediante l'interazione con il trasportatore associato al
processamento dell'antigene e l'inibizione del trasporto dei peptidi nel
reticolo endoplasmatico (York et al.,1994). In tal modo HSV può evitare o
limitare l'eliminazione da parte dei CTL (v. il saggio di E.J.H.J. Wiertz, nel
II volume).
Cloni di cellule T CD4+ isolati dalle lesioni da HSV
sono specifici per molti antigeni virali, comprese l'abbondante componente del
tegumento VPl6 e le glicoproteine dell'involucro gB, gC e gD. I ruoli diversi
delle cellule THl e TH2 sono stati studiati (Thomas e Rouse, 1997) nei modelli
murini della cheratite stromale erpetica (HSK, Herpetic Stromal Keratitis).
L'infezione oculare da HSV è la principale causa infettiva di cecità nei paesi
sviluppati; si pensa che HSK sia un'immunopatologia mediata dalle cellule T per
due ragioni: animali deficitari di cellule T risultano protetti dalla malattia;
HSK si sviluppa dopo che il titolo virale sta già diminuendo, quando le
risposte T -specifiche contro HSV si stanno sviluppando nella cornea.
Poiché fattori genetici correlati al ceppo murino e al
ceppo virale sono coinvolti in HSK, non sorprende che differenti cellule
effettrici sembrino causare HSK in diversi sistemi modello. Nei topi BALB/c
infettati con i ceppi KOS o RE di HSV-l, HSK è mediata dalle cellule CD4+ che
secernono TNF e IFN-γ, ma in topi AlJ gli effettori di HSK sono i CTL CDS+.
Anticorpi anti-IFN-γ bloccano HSK, mentre l'infiltrazione della cornea da parte
delle cellule di Langerhans (come effetto dell 'infiammazione) è richiesta per
HSK, poiché MHC di classe II non è espresso dalle cellule corneali. Sono stati
descritti altri meccanismi che coinvolgono danni prodotti dagli anticorpi e dal
complemento e una forma di citotossicità cellulare mediata dagli anticorpi e
dalle cellule NK. Per evidenziare la complessità di questi meccanismi, HSK può
essere indotta anche da un clone CD4+, mantenuto a lungo in coltura, specifico
per gD che produce IL-4, cioè un clone TH2.
Non esiste nessun vaccino efficace contro HSV. Gli
anticorpi neutralizzanti specifici per le glicoproteine dell'involucro gD e gB
possono proteggere e migliorare l'affezione ricorrente nel modello della cavia
e queste glicoproteine sono candidate per lo sviluppo di un vaccino. Altri
approcci promettenti comprendono i virus vivi attenuati o geneticamente resi
meno efficienti e l'immunizzazione mediante DNA. Un vaccino che prevenga
completamente le infezioni da HSV potrebbe essere irraggiungibile, ma
l'induzione combinata di anticorpi e immunità cellulare a livello delle mucose
potrebbe limitare la replicazione iniziale di HSV e l'instaurarsi
dell'infezione latente, portando a una forma più lieve o asintomatica
dell'affezione ricorrente.
Conclusioni
La risposta immunitaria è essenziale nella difesa
contro le infezioni virali, ma è anche in grado di danneggiare l'ospite. Senza risposte
immunitarie, i virus si diffonderebbero in modo incontrollato il che, si
assume, sarebbe dannoso per l'ospite. Quando vengono sintetizzate le proteine
virali, queste riducono la capacità della cellula di sintetizzare le sue
normali proteine. È da notare tuttavia che alcuni virus non danneggiano
particolarmente le cellule in assenza di risposta immunitaria. Si può sostenere
che questa possa essere buona o cattiva non solo per l'ospite, ma anche per il
virus. L'infiammazione che si genera dalla risposta immunitaria è spesso
d'aiuto al virus per raggiungere un nuovo ospite e, poiché i virus sopravvivono
solo se trasmessi a nuovi ospiti, la risposta è un aiuto essenziale per molti
virus.
Bibliografia citata
ALWAN, W.H., KOZLOWSKA, W.l,
OPENSHAW, P.l.M. (1994) Distinct types of lung disease caused by functional
subsets of antiviral T cells. J. Exp. Med., 179, 81-89.
AUTRAN, B., CARCELAIN, G., LI,
T.S., BLANC, C., MATHEz, D., TUBIANA, R., KATLAMA, C., DEBRE, P., LEIBOWITCH,
J. (1997) Positive effects of combined antiretroviral therapy on CD4+ T cell
homeostasis and function in advanced HIV disease. Science, 277, 112-116.
BACHMANN, M.F., ZINKERNAGEL,
R.M. (1996) The influence of virus structure on antibody responses and virus
serotype formation. Immunol. Today, 17, 553-558.
BIRON, C.A. (1997) Activation
and function of natural killer cell responses during viraI infections. Curr.
Opin. Immunol., 9, 24-34.
DANIEL, M.D., KIRCHHOFF, F.,
CZAJAK, S.C., SEHGAL, P.K., DEsROSIERS, R.C. (1992) Protective effects of a
live attenuated SIV vaccine with a deletion in the nef gene. Science, 258,
1938-1941.
DENNY, F.W. J.R. (1995) The
clinical impact ofhuman respiratory virus infections. Am. J. Respir. Crit.
Care. Med., 152 (4) Suppl., S4-S12.
FIORENTINO, D.F., BOND, M.W.,
MOSMANN, T.R. (1989) Two types of mouse T helper cell (IV): T H2 clones secrete
a factor that inhibits cytokine production by THl clones. J. Exp. Med., 170,
2081-2095.
HEILMAN, C.A. (1990)
Respiratory syncytial and parainfluenza viruses. J. Infect. Dis., 161, 402-406.
HILL, C.M., DENG, H., UNUTMAZ,
D., KEWALRAMANI, V.N., BASTIANI, L., GORNY, M.K., ZOLLA PAZNER, P.S., LITTMAN,
D.R. (1997) Envelope glycoproteins from human immunodeficiency virus types 1
and 2 and simian immunodeficiency virus can use human CCR5 as a coreceptor for
viraI entry and make direct CD4-dependent interactions with this chemokine
receptor. J. Virol., 71, 6296-6304.
HO, D.D., NEUMANN, A.U.,
PERELSON, A.S., CHEN, W., LEONARD, lM., MARKOWITZ, M. (1995) Rapid tumover of
plasma virions and CD4 Iymphocytes in HIV-l infection. Nature, 373, 123-126.
LU, R., PAXTON, W.A., CHOE, S.,
CERADINI, D., MARTIN, S.R., HORUK, R., McDoNALD, M.E., STUHLMANN, H., KOUP,
R.A., LANDAU, N.R. (1996) Homozygous defect in HIV-l coreceptor accounts for
resistance of some multiply-exposed individuals to HIV-l infection. CelI, 86,
367-377.
MOSMANN, T.R., SAD, S. (1996)
The expanding universe of T cell subsets: Thl, Th2 and more. Immunol. Today,
17, 138-146.
MOSS, P.A., ROWLAN-JONES, S.L.,
FRODSHAM, P.M., McADAM, S., GIANGRANDE, P., McMICHAEL, AJ., BELL, lI. (1995)
Persistent high frequency of human immunodeficiency virus-specific cytotoxic T
cells in peripheral blood of infected donors. Proc. Natl. Acad. Sci. USA, 92,
5773-5777.
NOWAK, M.A et al. (1995) Antigenic
oscillations and shifting immunodominance in HIV-l infections. Nature, 375,
606-611.
OPENSHAW, P.J.M. (1995)
Immunopathological mechanisms in respiratory syncytial virus disease. Seminars
in immunopathol., 17, 187-201.
PEREIRA, R.A, TSCHARKE, D.C.,
SIMMONS, A (1994) Upregulation of class I major histocompatibility complex gene
expression in primary sensory neurons, satellite cells, and Schwaun cells of
mice in response to acute but not latent herpes simplex virus infection in vivo.
J. Exp. Med., 180, 841-850.
SIGURS, N., BJARNASON, R.,
SIGURBERGSSON, F., KIELLMAN, B., BJÖRKSTÉN, B. (1995) Asthma and immunoglobulin
E antibodies after respiratory syncytial virus bronchiolitis: a prospective
cohort study with matched controls. Pediatrics, 95, 500-505.
SIMMONS, A., TSCHARKE, D.C.
(1992) Anti-CD8 impairs clearance of herpes simplex virus from the nervous
system: implications for the fate of virally infected neurons. J. Exp. Med.,
175, 1337-1344.
STREET, N.E., SCHUMACHER, J.H.,
FONG, T.A., BASS, H., FIORENTINO, D.F., LEVERAH, J.A, MOSMANN, T.R. (1990).
Heterogeneity ofmouse helper T cells. Evidence from bulk cultures and limiting
dilution cloning for precursors of T H 1 and T H2 cells. J. Immunol., 144, 1629-1639.
THOMAS, J., ROUSE, B.T. (1997)
Immunopathogenesis of herpetic ocular disease. Immunol. Res., 16, 375-386.
YORK, L.A., ROOP, C., ANDREWS,
D.W., RIDDELL, S.R., GRAHAM, F.L., JOHNSON, D.C. (1994) A cytosolic herpes
simplex virus protein inhibits antigen presentation to CD8+ T Iymphocytes.
CelI, 77, 525-535.
Bibliografia generale
FIELDS, B.N., KNIPE, D.M.,
HOWLEY, P.M., CHANOCK, R.M., MELNICK, lL., MONATH, T.P., ROIZMAN, B., STRAUS,
S.E., a c. di, FieIds viroIogy, 3' ed., Filadelfia, Lippincott-Raven, 1996.
JANEWAY, C.A. JR., TRAVERS, P.
Immunobiology, 3' ed., Londra-S. Francisco, Current Biology, e New Y ork,
Garland Publishing Inc., 1997.
MIMS, C.A. Mims' pathogenesis
of infectious disease, 4' ed., Londra, Academic Press, 1995.
PAUL, W.E., a c. di,
Fundamental immunology, 3' ed., New York, Raven Press, 1993 “
(fonte: http://www.treccani.it/enciclopedia/difese-contro-le-infezioni-virali_%28Frontiere-della-Vita%29/
)
------------
da ciò si evince, che forse il peggior virus della Terra
sia proprio l’uomo…o colui che definiamo tale ma che non lo è, io
rifletterei proprio su questa deduzione, per comprendere che l’Umanità si trova
ad un bivio, che non può più volgere lo sguardo da un’altra parte, ma deve
ricordare il suo Progetto per cui sta qui su questa Terra e
concretizzarlo… questo periodo di stop, in definitiva, era indispensabile…ascoltate
il vostro Cu♥re nel silenzio della vostra casa che vi sussurrerà di voi… Buon
ritorno a voi, Gaia vi aspetta!
a
cura di Maria Caterina Ranieri

{kind=link}
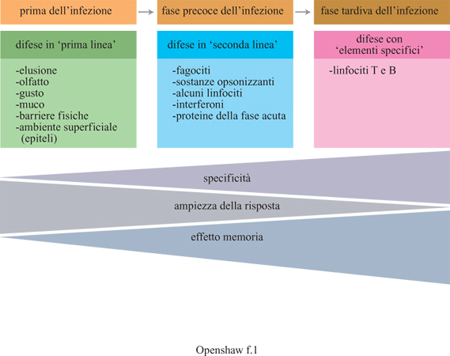{kind=link}
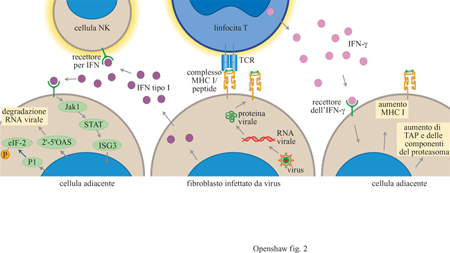{kind=link}
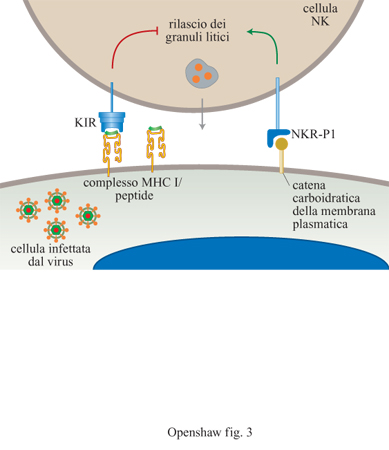{kind=link}
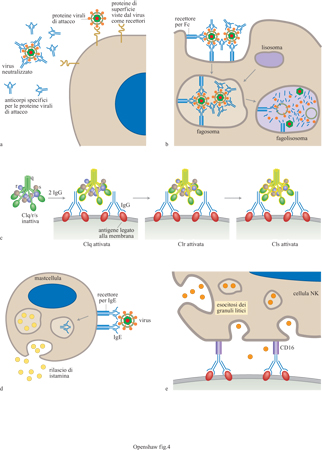{kind=link}
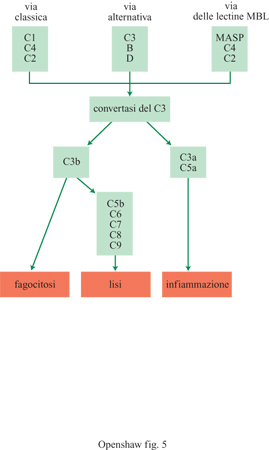{kind=link}
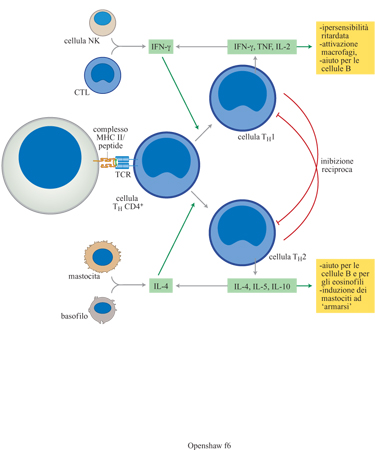{kind=link}
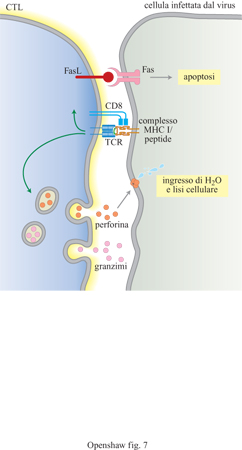{kind=link}
Nessun commento:
Posta un commento